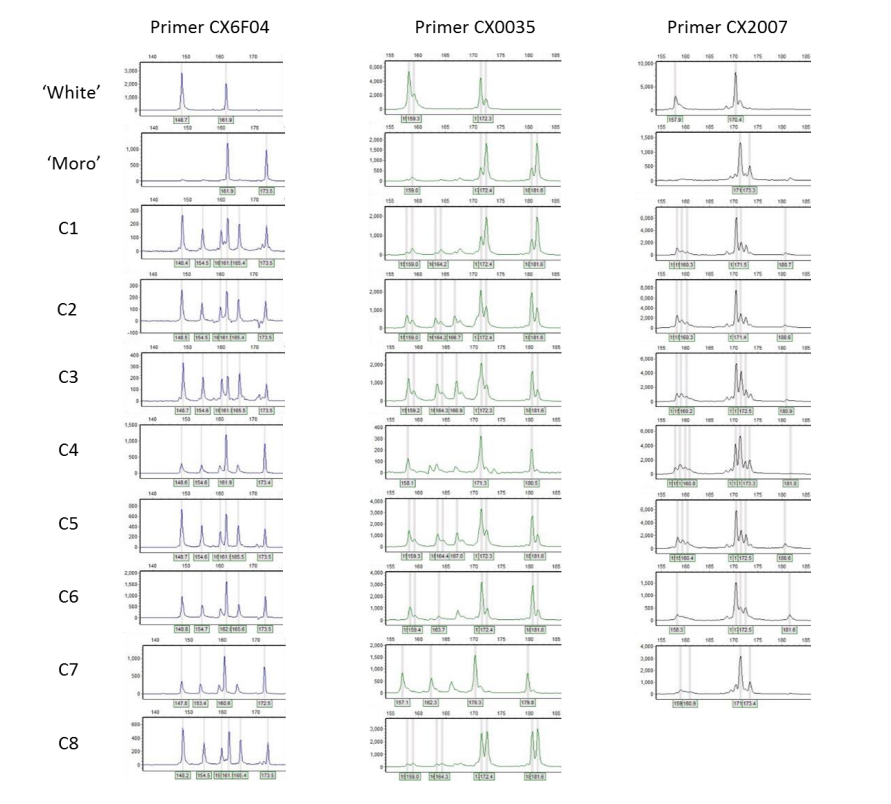
This is a valuable account on man-made periclinal chimeras between apple varieties. Thank you to quinoah on the Tropical Fruit Forum for sharing the original text.

187 SCHOLLE BOOKLET
Court Councilor Josef Loschnig
SCHOLLE-VERLAG WIEN I STUBENRING 6 (Scholle Publishing House, Vienna I, Stubenring 6)

Scholle Library, 187th Booklet
Improvement of Fruit Varieties
Hybrids, Mutations, Twin Fruits, Chimeras, Xenia
The Findings of Luther Burbank and J. W. Michurin
By Josef Löschnig
Former Lower Austrian State Director of Viticulture and Pomiculture (Fruit Growing)
With 41 illustrations
1948
Scholle-Verlag, Book Trading Company, Ltd., Vienna I., Stubenring 6
W 140, Lö 1042, Blanche von Melrose + Deans Codlin + Titowka.
Four-variety chimeras:
A 3, Lö 993, Charlamowsky + Kassler Renette + Peasgood Goldrenette + Adersleber Kalvill,
W 59/85, Lö 1037, (Jonathan + Jakob Lebel) + (Jonathan London Pepping).
A 1, Lö 1000, Fiessers Erstling + Kaiser Alexander + Gelber Bellefleur + Kaiser Alexander,
A 8, Lö 1001, Kaiser Alexander + Schöner von Boskoop + Großer Wellington + Bohnapfel
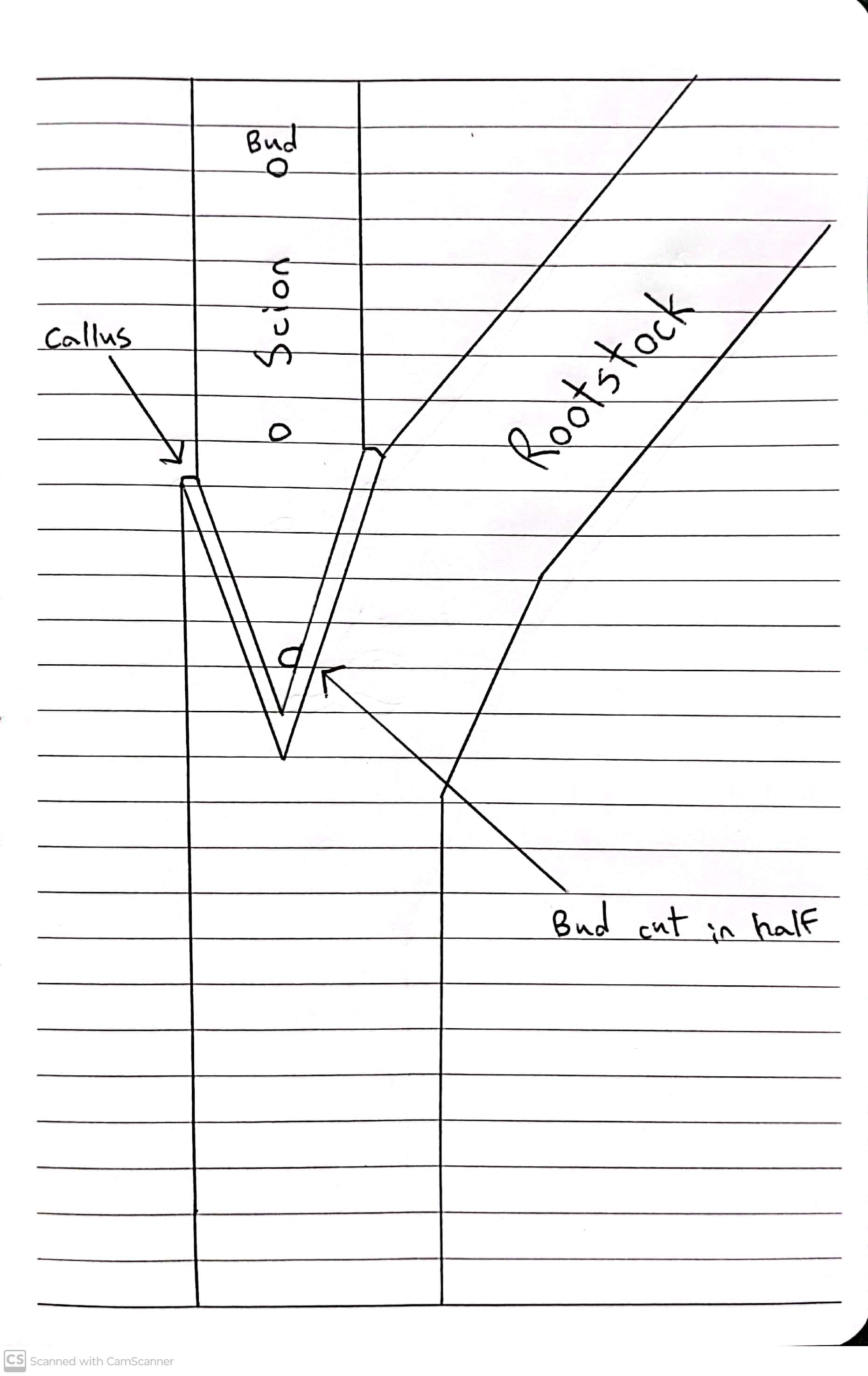
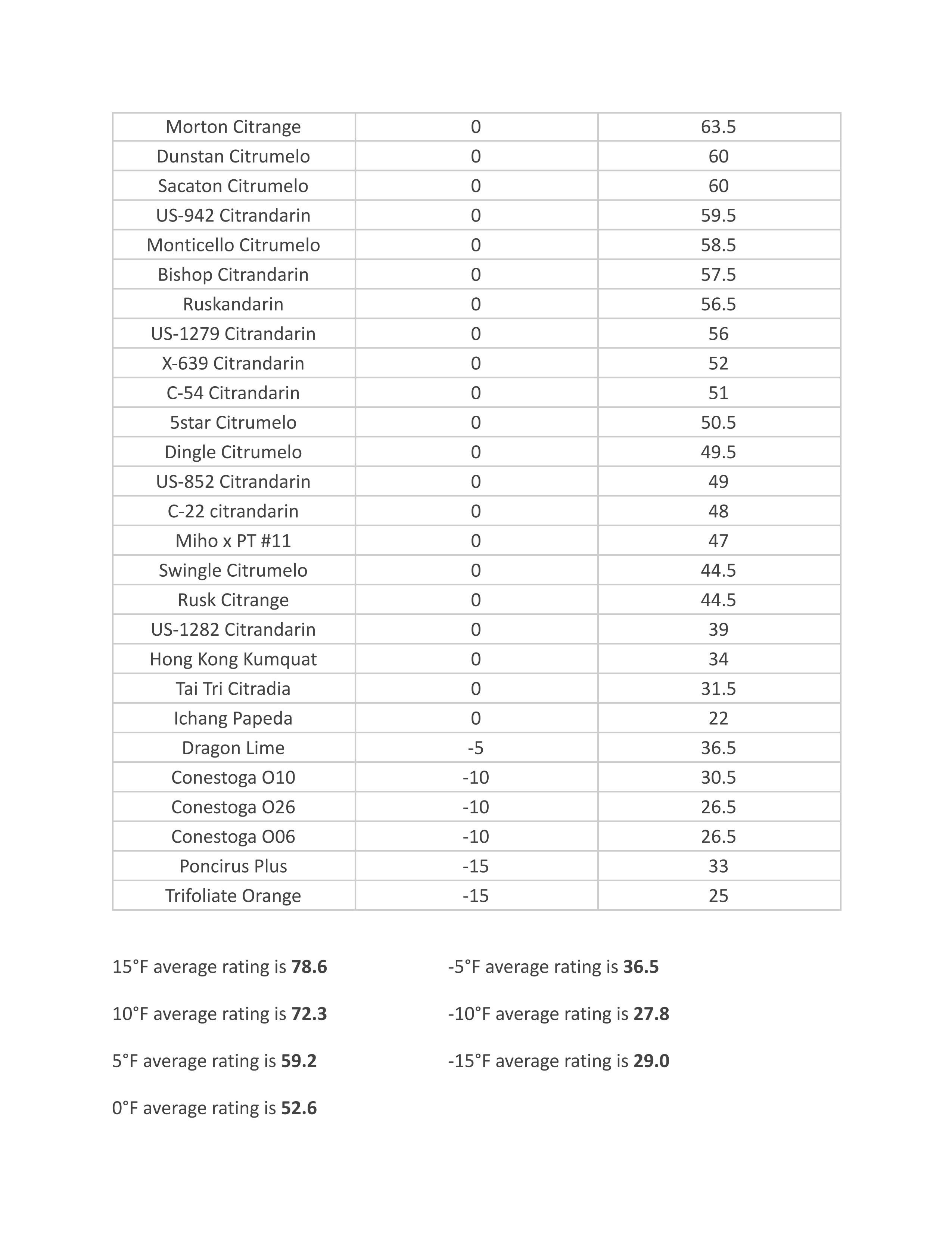
Fig. 12. Shoot-tip grafting according to Löschnig.
1 Trimmed shoot tips placed against one another;
2 Shoot tip showing the direction of the cut;
3 Fused graft after budding (without leaves);
4 and 5 Enlarged view of the shoot-tip graft in longitudinal section.

g) Findings to date on graft-chimeras
Experience with graft-chimeras is not yet sufficient to form a definitive judgment. Based on findings to date, which are supported by my work and the work of Wszolek, the following can be said in summary:
Through shoot-tip grafting according to Löschnig, graft-chimeras (“fable varieties”) can be achieved in apples and pears, and predictably also in stone fruit species. With correct execution, a successful fusion rate of 10% to 20% can be expected. The combination of varieties is best indicated by a diagram, such as those included in Fig. 6 and 10.
In shoot-tip grafting, the growth tips (vegetation cones) of two terminal buds—in which growth energy is at its highest—are united. Terminal buds of both leading and side shoots can be used for grafting. Shoot-tip grafting represents a layering (ablactation) of the shoot tips of two…
Fig. 13. Shoot-tip grafting (1) of two side shoots on an espalier wall in Wszolek’s garden. (Photographed by: Dr. Duhan.)

Budding occurs from the contact surfaces of the two growth tips, i.e., in the center or to the side of the graft; it therefore receives genetic traits from both parts, leading to the development of a mixture of tissues from two varieties. Often, side shoots also emerge from the accessory buds, but these must be suppressed, Fig. 13 and 14.
The chimeric graft shoot is usually weak, highly prone to fruit formation, and often does not reach full development until the second year. In the 2nd to 3rd year, you can already harvest the first fruits. When grafting buds that already have blossom buds, fruits can be obtained as early as the year of the graft. Fig. 15.
Fig. 15. Shoot-tip grafting in the summer of the first year. The new growth has already set a flower bud. (Wszolek’s garden.)
The first fruits are highly variable, often large to oversized, but also small and deformed. Based on Wszolek’s current experience, it can be assumed that during the cytoplasmic mixing via shoot-tip grafting, individual varieties show a certain dominance (graft dominance), e.g., Winter Gold Pearmain, Roter Eiserapfel, etc. The fruits of the second and third years are more uniform. Whether they can be kept completely uniform and stable will be shown by further ongoing experiments involving grafting onto various rootstocks.

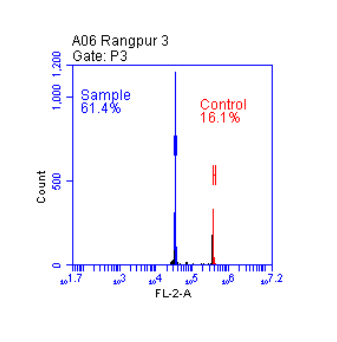
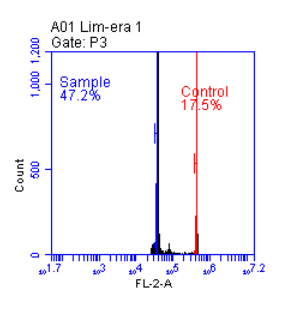
The chimeric character of the fruit manifests in fruit size, shape, and color, in taste, in the chemical composition of the flesh, and particularly through the appearance of “chimera marks.”
The chimera mark usually consists of a brown, wide stripe or one or more such spots on the skin, which can likely be attributed to superficial wound-cork formation. This is probably a reaction to the forced mixing of the fruit flesh tissues of two varieties. The chimera mark does not possess the color of either of the two varieties, but is fairly uniform across all chimeras, only slightly intermixed with the color of one or the other variety.
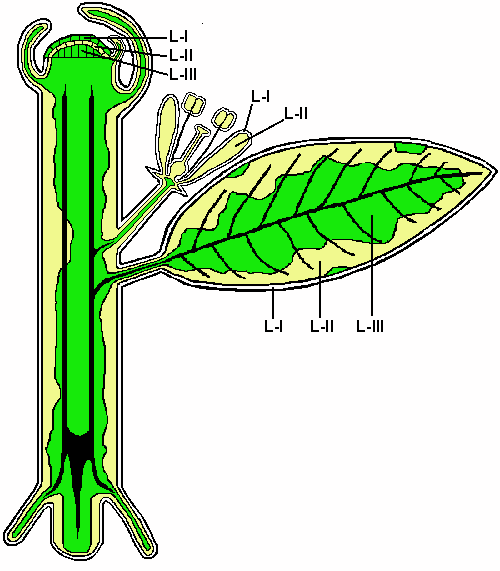
Fig. 16. Leaves of the graft-chimera Edelstein Delicious + Goldparmäne, shown at half size from the same tree.
1 Edelstein Delicious,
2 Goldparmäne,
3 Chimeric leaf. (Wszolek’s garden.)

…mixed into all chimeras, just slightly tinged by the color of one or the other variety.
The flesh of the fruits varies greatly from one to another, yet within a single fruit, it is fairly uniform with no visible boundary, even though such a boundary would be expected from the mixing of tissues from two varieties. In the Eiserapfel + Cellini chimera, the flesh beneath the chimera mark had a 2% higher sugar content than the remaining parts of the fruit flesh.
The seed of the chimera is unbelievably diverse. In isolated cases, one finds seeds that resemble one or the other parent variety and are normally developed. Mostly, the seeds are stunted and hollow; if individual seeds are present, they display a white tip or a white spot. The germination capacity of the seeds has not yet been investigated, but is unlikely to exist.
The seed cavities are consistently smaller than in the parent varieties, which is due to the lack of seeds or hollow seeds. In several cases (about 12%), the seed cavities were filled with a thick, fleshy mass. In one case, Kaiser Alexander + Jakob Lebel, a displaced seed cavity was found in the fruit flesh attached to a vascular bundle.
Most chimeric fruits possess double vascular bundles running over one another. Sometimes the doubling of the vascular bundles is only partially, unilaterally, or locally noticeable.
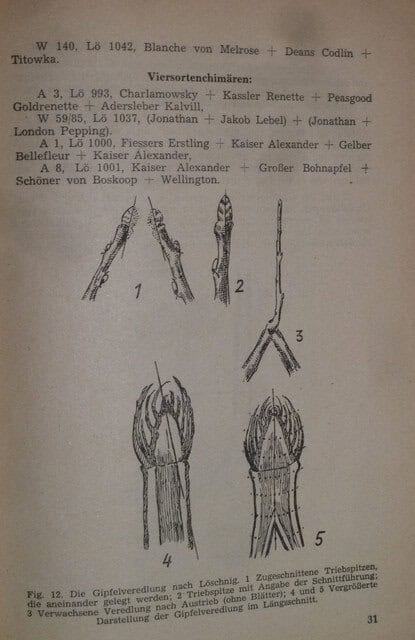
The leaves of the chimeras are formed fairly uniformly within the chimeric section, but show great differences compared to the parent varieties. In ten examined individual cases, the chimeric leaf was smaller than those of both parent varieties nine times, and significantly larger once. In isolated cases, the leaves are unequal and lean closer to those of the individual parent varieties, Fig. 16 and 17.
The taste of the chimeric fruits is very different compared to that of the parent varieties: often better, especially sweeter, sometimes worse. The sugar and acid content of the fruits of a chimera can vary by up to 2% for sugar and by 0.3% for acid. This variation is likely due on one hand to combinations of the tissue mixing, and on the other hand to the re-emergence of traits dormant in the parent varieties.
The status of the hereditary factors, the chromosomes, in chimeras has not yet been researched. One can assume two possibilities, as Professor Winkler established with the Tomato + Solanum nigrum (Nightshade) chimera:
a) The cell nuclei of all plant body cells of the chimera have the same chromosome number as the parent varieties; in apples and pears, diploid 34. In this case, it would be a mixing (not a fusion) of the tissues of the parent varieties.
b) The chimeras combine the chromosome numbers of the parent varieties in their body cells; in apples and pears, a diploid 34 + 34 = 68 chromosome set. In this case, it would have to be an asexual fusion of two plant cells. Winkler referred to these, which are characterized by particularly vigorous growth, as “burdons.”
Fig. 17. Leaves of the three-variety graft-chimera, shown at half size.
1 Bismarckapfel,
2 Baumanns Renette,
3 Berlepsch’ Renette,
4 Chimeric leaf. (Wszolek’s garden.)

In conclusion, we must state that today we do not yet have a full overview of the significance of graft-chimeras, as we do not know their subsequent behavior during vegetative reproduction via grafting.
It could be an incredibly interesting breeding innovation that produces new, even valuable plant forms, but which has no great economic significance due to its low stability.
However, it could also be possible that graft-chimeras have opened up a new way to improve our fruit varieties. At present, we know that through shoot-tip grafting we can achieve new forms with fabulous fruits and combine different characteristics of the varieties; if these remain stable, then we can also propagate them through grafting and have rendered a service to fruit growing.
Xenia (from the Greek for “guest gifts”) refers to all those changes in fruits, seeds, and other parts of a maternal plant that can be attributed to the male pollen during cross-fertilization (hybridization). In the case of pome and stone fruit, we are dealing with accessory fruits, which consist of the outer parts of the flower—the thick, fleshy calyx part—and enclose the seed. The outer floral and fruit parts do not participate in the formation of the seed. The fleshy part of the fruit therefore comes solely from the mother, while the seed comes from both parents as a result of the preceding fertilization. True xenia are therefore likely rare in pome and stone fruits. In several cases of observed xenia, subsequent experiments proved that these were due to other causes, mostly mutations. Dr. Duhan undertook extensive hybridizations in apples and examined the resulting fruits for xenia formation, without being able to detect any influence of the male pollen on the shape, color, sugar content, or pH value of the fruit. In practical fruit-growing circles, however, experiences that could be attributed to xenia formation are widespread.
In apricots, the stone shape is frequently altered by the cross-pollination partner. The Ambrosia apricot develops a slender stone with self-pollination, but a bulbous stone when fertilized by Breda or Hungarian Best. Due to the bulbous nature of the stone, the fruit shape is also altered: the fruit becomes wider and thicker. Here, the influence of the male pollen on the stone and fruit shape is obvious, even though this is a mechanical adaptation to the bulbous stone.